IUCN/SSC Otter Specialist Group Bulletin

|
©IUCN/SCC Otter Specialist Group Volume 28A Proceedings Xth International Otter Colloquium, Hwacheon, South Korea Citation: Evangelista, E. and Rosas, F.C.W., B. (2011a) The Home Range and Movements of Giant Otters (Pteronura brasiliensis) in the Xixuan Reserve, Roraima, Brazil . Proceedings of Xth International Otter Colloquium, IUCN Otter Spec. Group Bull. 28A: 31 - 37 The Home Range and Movements of Giant Otters (Pteronura brasiliensis) in the Xixuan Reserve, Roraima, Brazil Emanuela Evangelista1, Fernando C.W. Rosas2
1
Associação Amazônia NGO, Caixa Postal 1212 Ag. Saldanha Marinho, Manaus - AM, 69080-710, Brazil (Phone: +55 (92) 92165584, mobile: +39 347 1375202 e-mail: eman_evan@hotmail.com |

 |
|
Abstract: A population of Giant otter was studied in the central Brazilian Amazon at the Xixuaú Reserve, Roraima (0º48’S, 61º33’W), in order to gather preliminary information on spatial use of habitat. Through the period 2000–2003, 80 animals have been recorded in the study area, of which 9 solitary (11.2%) and 71 (88.7%) in groups of an average of 4.46 individuals (n=15; biggest group=9). For 4 Giant otter groups, dry season home ranges were estimated at approximately 8 km in average (n=6; minimum size 4.6 km, maximum size 10.5 km), along watercourses of 52.8 m of average width (n=8). Groups’ home ranges in the study area can overlap. An expansion in direction of flooded forest areas has been observed during the flooded season as well as a temporary contraction during the period of cub rearing. The periodic and complete counting of the animals allowed estimating the density of the species in the study area (1 animal/1km). |
| Keywords: habitat use; home range; territorial behavior; Pteronura brasiliensis; giant otter density |
| Française | Español |
INTRODUCTION
Giant otter is endemic to South America and endangered throughout its geographical distribution area (IUCN, 2006) due to habitat degradation and destruction (Carter and Rosas, 1997; Groenendijk et al., 2005). Although the species is threatened there is still limited understanding about its ecological requirements and movement patterns, its home range being among the lacking information (Utreras et al., 2005). With the aim of estimating the potential size of Giant otter home range and the ecological density of the species we studied the resident population of giant otter at the Xixuaú Reserve. The study of a species in absence of anthropogenic disturbance can provide helpful information for conservation strategy programs. The Xixuaú Reserve is a protected area of 178,000 ha, accessible exclusively by boat from the city of Manaus with a journey of approximately 500 km. The area is virtually uninhabited, with a human population density of 0.04 persons per km2 and hence consents the gathering of information in a pristine area.
MATERIAL AND METHODS
The Xixuaú Reserve (0º48’S, 61º33’W; altitude 30 m a.s.l.) is situated on the right hand bank of the Jauaperi River, which marks the border between the Brazilian states of Amazonas and Roraima. It consists mainly of primary tropical rainforest crossed by a black water river and many creeks. The yearly rainfall is about 2000 mm with a dry period from October to February. During the flooded season, the water level usually rises up to 10 m, flooding wide portions of the forest and sandy beaches along the watercourses.
The main study area consists of approximately 60 km2 of the Jauaperi river system, in correspondence of the Xixuaú creek (Fig. 1). In order to verify the existence of a buffer zone outside the core-investigated section, a wider area along 40 km of Jauaperi River were monitored.
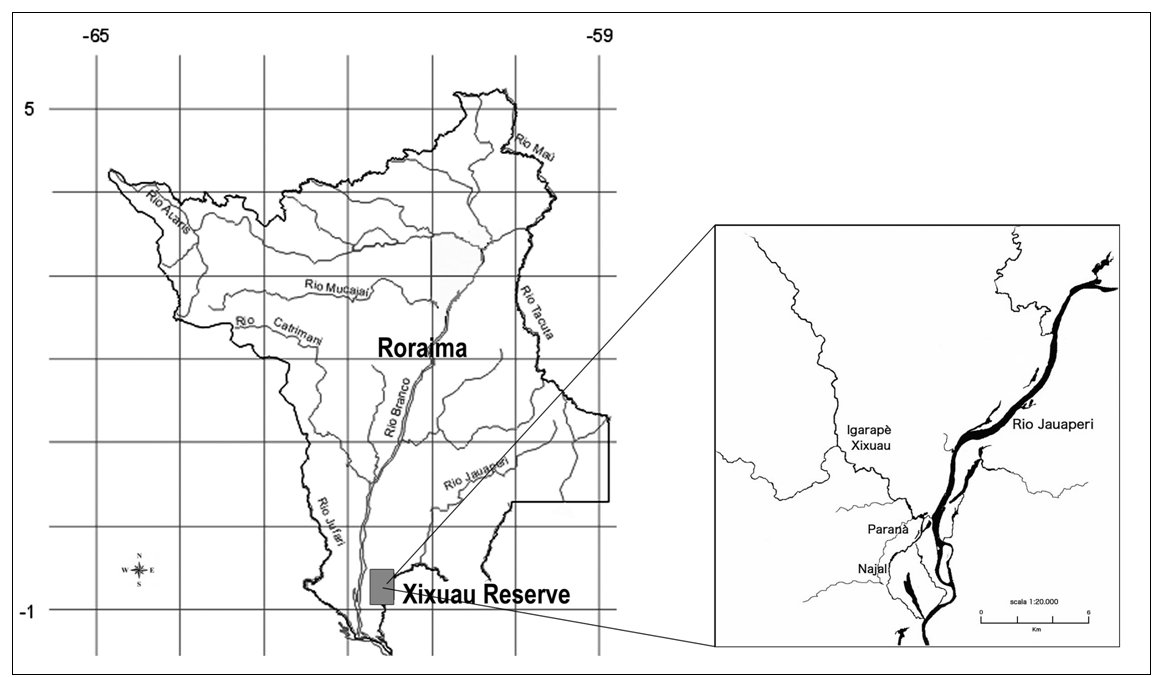 |
| Figure 1. Location of the study area. (click for larger version) |
Between October 2000 and March 2003, 14 months have been spent in the study area, carrying out daily surveys using a paddle-propelled canoe. Riverbanks were patrolled in order to record the geographical location (WGS 84) of indirect signs (i.e. latrines, campsites, dens, tracks) and direct observations of giant otters. A photographic catalogue of the throat pattern of the animals, aimed to avoid double counters, has been carried out using a Camcorder. The animals have been followed at a distance of 10–100 meters or observed from shelters. Dry season home range was estimated using a combination of direct observations and indirect signs of presence. Dens and campsites in recent use have been attributed to a family group based primarily on sights (i.e. the group was seen using the site). When the direct observation was not possible, the data collected in the same or previous/next day has been taken into account. That is, a site in recent use has only been attributed to an otter group if (1) the group was sighted in the same area, (2) the neighbour group was sighted in its own territory, and (3) no solitary animal was sighted. Based on the records along the watercourses, home range size has been estimated by considering only the linear distance. In order to allow future comparisons of the data some ecological characteristics of the river system have been collected (i.e. width, water depth, visibility of water measured by Secchi disk). The periodic and complete counting of the animals allowed the estimate of the species density in the study area.
RESULTS
Occurrence of Giant otter
Eighty animals have been sighted, 9 of them classified as solitary (11.2%) and 71 (88.7%) in groups. A total of 15 groups of Giant otters have been recorded, the greater composed by 9 animals, with an average size of 4.46 individuals.
Home range
It was possible to estimate the home range size of 4 groups (named M, BN, N and U) during the dry season (Tab. 1). The largest home range was 10.5 km (BN group) and the smallest was 4.6 km (M group) in linear distance, along watercourses of 52.8 m of average width (n=8). Exclusively for the M group the home range was calculated in three different dry seasons and it varied from 4.6 km to 9.7 km (Fig. 2, Fig. 3).
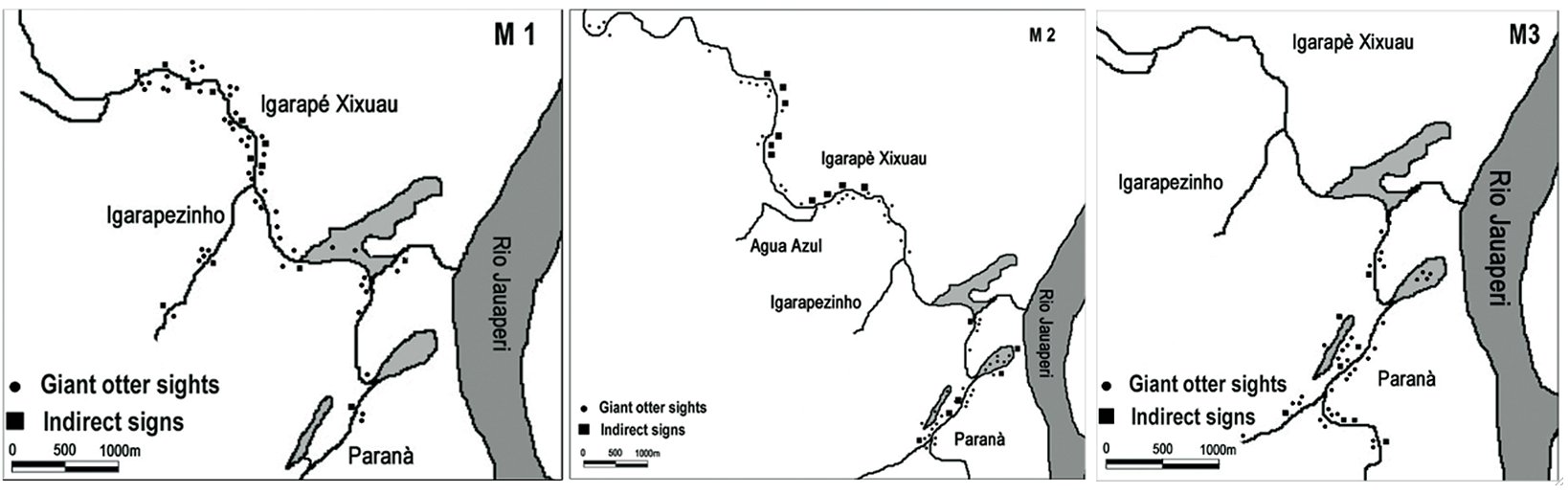 |
| Figure 2. Home range of the “M” Giant otter group estimated in the dry season 2000–2001, 2001–2002 and 2002–2003 (from left to right). (click for larger version) |
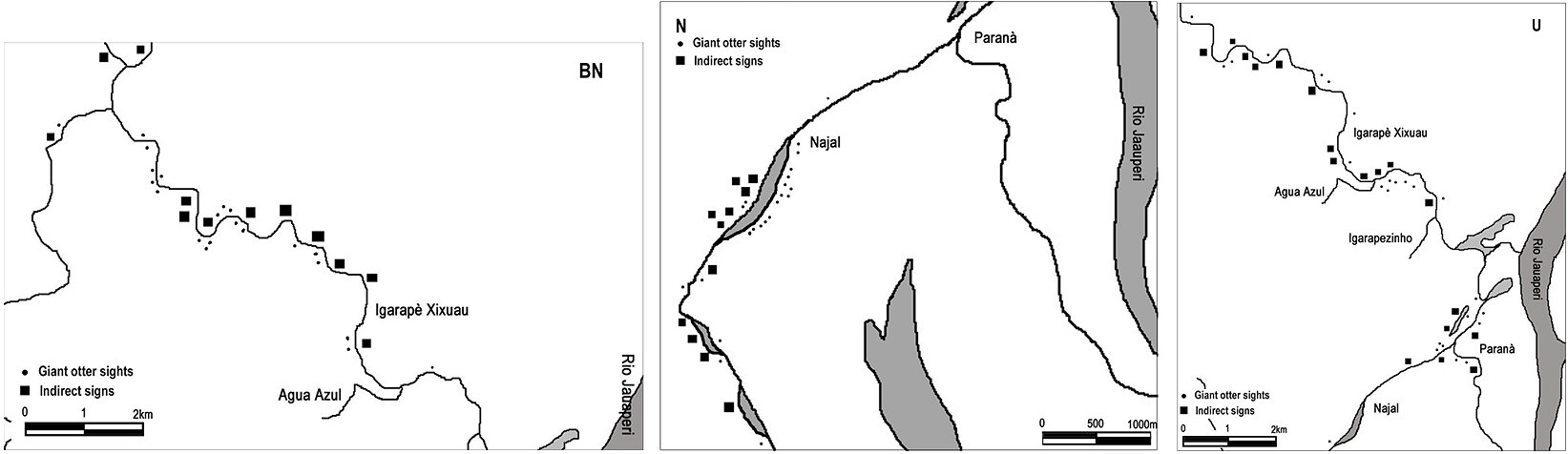 |
| Figure 3. “BN”, “N” and “U” groups home range estimates (from left to right).. (click for larger version) |
Unlike the M, BN and N groups that were sighted every year approx. in the same area, the U group appears in the study area only in the 2001–2002 dry season. The previous year it was known as resident in the buffer zone (União Lake, 15 km up the Jauaperi River), where it went back the next year. During 2001–2002 dry season the U group invaded mainly the M group territory, expanded also towards the BN and N home ranges, both using existing dens and campsites and building new ones. Agonistic behavior has not been recorded, as the groups seemed to avoid each other.
The differences in home range, did not seem to be related to the size of the Giant otter groups, which, for example in the 2nd season studied, had three individuals in the M group, six in both the BN and the U groups, and seven in the N group. Nevertheless, a temporary contraction has been observed in the M group during the period of cub rearing. In the first season 2000–2001 the group, composed by an alpha couple and a cub 0–3 months old, used a home range of 6.3 km; in the second season (2001–2002) the group, composed by two adults and one subadult, used a home range of 9.7 km. In the third season (2002–2003) the group, composed by 3 adults and 2 cubs 2.5–5.5 months old, reduce again the home range to 4.6 km.
The Xixuaú watercourse system consists of little streams and fluvial lakes; during the dry season the watercourses width ranges from 20 to 100 m, with 30–150 cm of water depth and few deeper pools of 200–400 cm deep. Visibility, measured by Secchi disk, ranges from 50 to 200 cm. During the 2002 flooded season, between May and July, was possible to observe giant otters only in 13 occasions, 7 of them unfruitful in recognizing the animals due to the dense vegetation of the flooded forest. Despite this, it was possible to record 26 sites in recent use (dens and campsites), located on emerged land under the flooded forest or in seasonal watercourses temporarily accessible, all of them characterized by little width (5–10 m), good water depth (4 m) and high visibility (4 m). (Fig. 4).
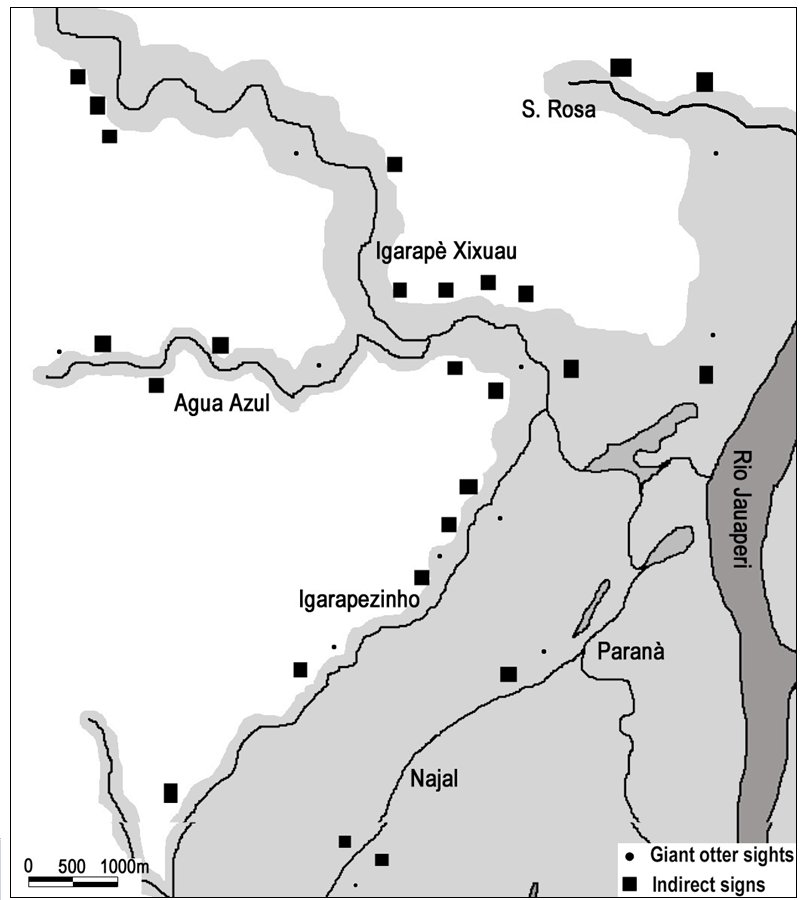 |
| Figure 4. Giant otter sights and indirect signs recorded during the flooded season 2002. It is possible to note the seasonal streams named Igarapezinho, Agua Azul, S. Rosa. (click for larger version) |
Density
The crude density calculated in the main study area, including land sections not used by the Giant otter, is 1 animal every 2 km2. The ecological density, considering only the habitat utilized by the species and not including the transient animals, is 1 animal per 1 km.
DISCUSSION
Home range is defined as the surface area regularly used by an individual or a social group for feeding, reproduction and parenting (Burt, 1943; Laidler, 1984). It should be defined with regard to a specific unit of time (Spagnesi and Randi, 1995) and should be analyzed with statistical methods in order to allow inter and intra-specific comparisons and to provide information on the intensity of the habitat use (Pedrotti et al., 1995). The method here utilized offers a spatial representation of the vital areas used by giant otter groups based on deductive analyses and does not allow a statistical definition. Moreover, it does not provide information on the habitat preferences and could supply an underestimation of the effective vital area of the species. Results must be interpreted as an estimate of the minimal area occupied by a single group, that is where they spend the main part of their time, reproduce, and are sighted more frequently.
Information on vital areas of Giant otters comes from studies carried out in Suriname (Duplaix, 1980), Guiana (Laidler, 1984), Peru (Schenk, 1999; Staib, 2005) and Ecuador (Utreras et al., 2005).
Although comparison of data is not always possible due to the use of different units of measure (km, km2), the results of the present work are within the range of values reported by other authors. For example, Duplaix (1980) observed 4–5 groups in Suriname distributed along 12 km of river including its affluent (Kaboeri Creek, width = 20 m). Home ranges overlapped and the territorial behavior of the animals was limited to a minor area of 2–3 km of river. In Guiana, Laidler (1984) calculated for 4 family groups a home range of 32 km of river, which overlapped and connected territories of 9.6 km. In Ecuador, if one considers only the linear distance along the river, Utreras (2005) found that home ranges of family groups varied from 8 to 19 km. It is probable that some habitat characteristics (i.e. hydrologic regime, food availability, etc.) as well as the species density in the area are responsible for the different values reported.
The home range size here presented is based on dry season movements of the giant otter groups. During the flooded season in the study area the water level rises up to 10 m, submerging all dens and campsites previously used by the groups. In this time of the year the data revealed a movement of the groups towards the flooded forest where fish disperses, as already reported by Duplaix (1980), who stated that the groups abandoned the former territories during the high water period. However, the observation of new sites located in correspondence of the dry season areas, suggests that in the study area an expansion of home range, more than an abandonment of the same territories, can take place.
In agreement with the reports from Ecuador (Utreras et al., 2005) the home range size at Xixuaú does not seems to correlate with the group size. Nevertheless, it is possible, as already mentioned by Laidler (1984), that increase or reduction of territories, like the temporary changes recorded in the M group, are linked to the development stages of the litter. In agreement with Duplaix (1980), Laidler (1984) and Utreras (2005) but contrasting with reports from Schenck (1999) and Staib (2005) in Peru, home range in the study area can partially overlap. However, each group has been regularly observed in a specific locality and seldom observed in different areas. The U group represents the single exception. Staib (2005) observed the change of a residential group in a cochas in Peru, during two successive census seasons. As the dynamic of the substitution was not known she assumed that the area was vacant when the new group occupied it. A sliding of the territories has also been documented by Duplaix (1980) who assumed that in her study in Suriname the new entry of the KI alpha couple was due to the contraction of the H group territories after the death of the reproductive female and her litter. In both cases the entering of a new group seems to be connected to the favorable condition of an available area. To our knowledge, such attempt of the U group of occupying a full area has never been recorded before and it could be connected to the high anthropogenic pressure that the area suffered in the 2001–2002. Commercial fishermen in the study area are known for strongly persecuting giant otters, their presence in the União Lake forced the inhabitants of the Xixuaú Reserve to create a new settlement in that lake in order to stop the invasions and protect the area. Afterwards, the U group went back to its former territory.
A number of authors calculated the ecological density for giant otters in their study areas. Our results do not differ very much from the data presented by Duplaix (1980) who reported in Kaboeri Creek a density of 1 animal for 0.5 km, although it is considerably higher than values calculated by Laidler (1984) in Guiana and Schenck (1999) in Peru (1 animal each 5.6 and 5.7 km, respectively). In different locations along its distribution the giant otter seems to prefer slow-flowing clear or black water creeks and rivers with good forest cover (Duplaix, 1980, 2002; Laidler, 1984; Rosas et al., 1999; van Damme, 2002). Hence, it is possible that the high density calculated for the Xixuaú river system could be biased as the main study area has been chosen between the preferred habitats. However, comparing the study areas, it is also possible to assume that it depends on the low anthropogenic pressure registered in a remote and protected environment such as the Xixuaú Reserve.
ACKNOWLEDGMENTS - We are in debt to the inhabitants of the Xixuaú Reserve and to the Associação Amazônia NGO for the logistic support. Special thanks are due to the native guides Carlos Alberto Nascimento and Alexandre Soares for their important help during the fieldwork. We also extend our gratitude to Christopher J. Clark who revised the English version of this manuscript.
REFERENCES
Burt, W.H. (1943). Territoriality and home-range concepts applied to small mammals. J. Mammal. 24: 346-352.
Carter, S.K., Rosas, F.C.W. (1997). Biology and conservation of the giant otter Pteronura brasiliensis. Mammal Rev. 27: 1-26.
Duplaix, N. (1980). Observations on the ecology and behavior of the giant river otter Pteronura brasiliensis in Surinam. Revue Ecologique (Terre Vie) 34: 495-620.
Duplaix, N. (2002). Guianas Rapid River Bio-assessments and Giant Otter Conservation Project - WWF Report.
[http://www.giantotterresearch.com/articles/WWF_Giant_Otter_Report_PDFMini.pdf accessed 13 July 2011]
Groenendijk, J., Hajek, F., Duplaix, N., Reuther, C., van Damme, P., Schenk, C., Stain, E., Wallace, R., Waldemarin, H., Notin, R., Marmontel, M., Rosas, F., Mattos de, G.E., Evangelista, E., Utreras, V., Lasso, G., Jaques, H., Matos, K., Roopsind, I., Botello, J.C. (2005). Surveying and monitoring distribution and population trends of the giant otter (Pteronura brasiliensis). Habitat 16: 1-500.
IUCN (2006). 2004 IUCN Red List of Threatened Species.
[www.iucnredlist.org. accessed 23 August 2007]
Laidler, L. (1984). The behavioral ecology of the giant otter in Guyana. Doctoral Dissertation, Univ. of Cambridge 1984; 319 pp.
Pedrotti, L., Tosi, G., Facoetti, R., Piccinini, S. (1995). Organizzazione di uno studio mediante radio-tracking e analisi degli Home Range: applicazione agli ungulati alpini. In: Spagnesi, M. e Randi, E. (eds): Applicazioni del radio-tracking per lo studio e la conservazione dei vertebrati. Suppl. Ric. Biol. Selvaggina, XXIII: 3-100
Rosas, F.C.W., Zuanon, J.A.S., Carter, S.K. (1999). Feeding Ecology of the Giant Otter, Pteronura brasiliensis. Biotropica 31(3): 502-506
Schenck, C. (1999). Lobo de Rio (Pteronura brasiliensis). Presencia, uso del habitat y proteccion en el Peru. GTZ/ INRENA, Lima, Peru; Spanish translation of German PhD dissertation, Università di Monaco, 177 pp.
Spagnesi, M., Randi, E. (1995). Applicazioni del radio-tracking per lo studio e la conservazione dei vertebrati. Suppl. Ric. Biol. Selvaggina, XXIII: 1-186
Staib, E. (2005). Eco-etologia del Lobo de Rio in Peru. Spanish translation of German PhD dissertation: Öko-Ethologie von Riesenottern (Pteronura brasiliensis) in Peru. Munich University, Germany, 195 pp.
Utreras, B.V., Suárez, R.E., Zapato-Ríos, G., Lasso, G., Pinos, L. (2005). Dry and rainy season estimations of giant otter, Pteronura brasiliensis, home range in the Yasuní National Park, Ecuador. LAJAM 4(2): July/December 2005 - ISSN 1676-7497
Van Damme, P.A., Wallace, R., Swaenepoel, K., Painter, L., Ten, S., Taber, A., Gonzales, J., Saravia, I., Fraser, A., Vargas, J. (2002). Distribution and population status of the giant otter Pteronura brasiliensis in Bolivia. IUCN Otter Spec. Group Bull. 19(2) : 2002
Résumé : Domaines Vitaux Et Déplacements Des Loutres Géantes (Pteronura brasiliensis) Dans La Reserve De Xixuau, Roraima, Brésil
Une population de loutres géantes a été étudiée dans le centre de l'Amazonie brésilienne dans la réserve Xixuaú (Roraima), dans le but de recueillir des données écologiques et comportementales. Durant la période 2000-2003, 14 portées ont été enregistrées, toutes entrent les mois d’Août et Décembre au cours de la décrue et des périodes de basses eaux. Le pic de reproduction semble être dépendant de la disponibilité en nourriture dans le système fluvial. La taille des portées varie de 1 à 4 petits avec une moyenne de 1,8 (n=12). Il a été confirmé qu'une forte corrélation positive existe entre la taille du groupe et le taux de reproduction, mais aucune preuve de "baby-sitting» décrit par d'autres auteurs n'a été trouvée. L'étude fournit des informations sur le développement des loutrons et confirme que tous les membres du groupe sont impliqués dans l'élevage des petits.
Revenez au dessus